
 下载专区
下载专区 English
English科研进展

亮文解读 | 生物调控产酶溶杆菌OH11产生新的WAP-8294A环脂缩肽
Sustainable and Environment-Friendly Agricultural Pest Management
· 第十篇 ·
▎论文ID
发表年份:2022年
 : ldu3@unl.edu
: ldu3@unl.eduCite this article :
Jing ZHU, Yuan CHEN, Liangcheng DU. PRODUCTION OF NEWWAP-8294A CYCLODEPSIPEPTIDES BY THE BIOLOGICAL CONTROL AGENT LYSOBACTER ENZYMOGENES OH11. Front. Agr. Sci. Eng., 2022, 9(1): 120‒132 https://doi.org/10.15302/J-FASE-2021410
1. 构建了产酶溶杆菌突变体,以生物合成WAP-8294A环脂缩肽。
2. 确定了生物合成WAP-8294A环脂缩肽的必要和非必要附属基因。
3. 利用超高效液相色谱-质谱联用技术鉴定了6种新的WAP-8294A类似物的结构。
4. 检测到了3种脱氧类似物,可支持ORF4的天冬酰胺羟基化功能推测。
由微生物产生的具有全新化学结构和独特作用位点的天然活性化合物已经逐渐成为对抗病原菌抗药性问题的重要方法。溶杆菌是一种广泛分布于土壤和水中的滑行革兰氏阴性细菌,其中多种溶杆菌可以大量产生胞外裂解酶,能够抑制病原菌的生长,有望成为新的具有生物活性的天然产物重要来源。从溶杆菌中已鉴定了包括多环特特拉姆酸大环内酰胺(PoTeM)、环脂缩肽、头孢型β-内酰胺和吩嗪等多种类型的活性天然产物。而从溶杆菌中分离的非核糖体肽如环肽tripropeptins, WAP-8294As, lysocins, WBP-29479A1 和lysobactin,含有复杂的结构并显示了较高的抗菌活性。其中WAP-8294A环肽家族能有效地抑制革兰氏阳性病原菌,该家族含有至少19个结构紧密相关的成员,主成分WAP-8294A2的结构已通过质谱、一维和二维核磁共振谱获得解析,而微量成分A1,A4,Ax8,Ax9和Ax13等通过质谱鉴定,其余成分结构尚未获得鉴定。
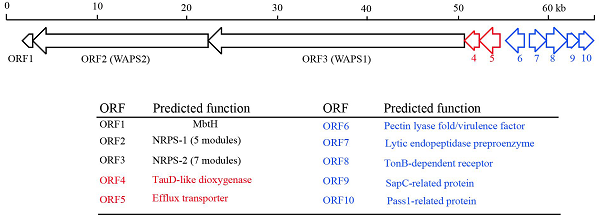
产酶溶杆菌OH11的WAP-8294As生物合成基因簇(BGC)包含三个结构基因ORFs 1–3,分别编码多模块非核糖体肽合成酶(NRPS)WAPS1和WAPS2以及NRPS相关蛋白MbtH(图1)。与三个结构基因聚集在一起的是七个辅助基因,ORFs 4–10,它们可能在WAP-8294As的修饰、调控和抗性中起作用。现阶段的研究主要集中在使用合成生物学等方法对WAP-8294A BGC进行改造提高已知化合物的产率,而几乎没有人尝试通过遗传学方法改造溶杆菌使其产生新的WAP-8294A类似物。本研究中,我们对WAP-8294A BGC辅助基因进行了一系列的基因操作,并在单点突变株中发现了WAP-8294As的脱氧类似物。此外,使用超高效液相色谱-高分辨率串联质谱(UHPLC-HR-MS/MS),我们检测到六种新的WAP-8294A化合物。这些结果表明,可以通过操纵生物合成基因的方法来扩大产酶溶杆菌OH11的生防化合物谱。
▎结果与分析
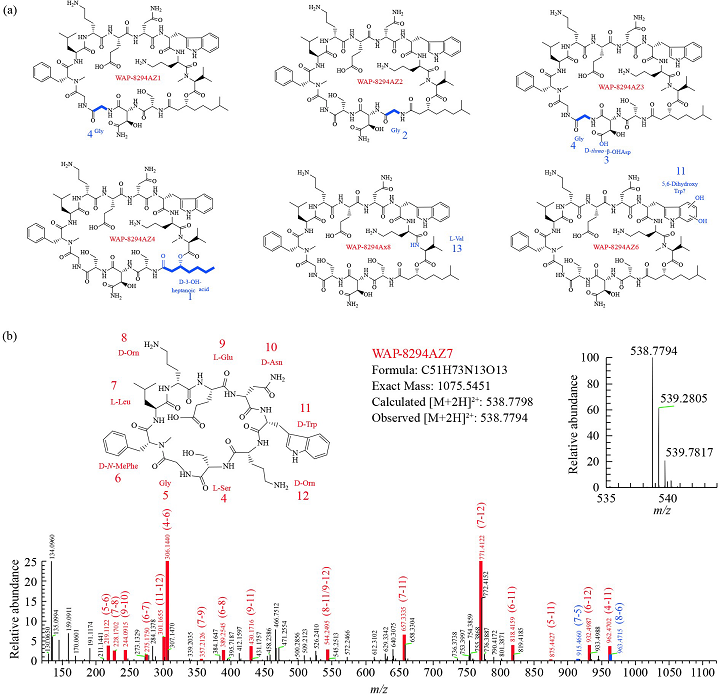
为了验证通过操纵与结构基因成簇的辅助基因(MbtH和NRPS的ORFs 1–3)可以产生新的WAP-8294A化合物的推测,我们对WAP基因簇中的基因进行了同框缺失突变(图1)。首先,我们分别敲除了ORFs6,7,9和10四个下游基因,并分析野生型和突变体的代谢产物,结果表明四个辅助基因对OH11产生WAP-8294A不是必需的。为了进一步证实这一点,我们同时敲除了ORF6到ORF10五个下游基因,产生了一个五联突变体,HPLC显示该五联突变体仍然能产生WAP-8294A化合物(图2),表明这五个下游基因对于WAP-8294A生物合成不是必需的。然后,我们验证了WAP基因簇中剩余的两个辅助基因,ORF4和ORF5,HPLC显示在单独敲除ORF4或ORF5的突变株OH11ΔWAP-ORF4或OH11ΔWAP-ORF5中没有产生WAP-8294A化合物,表明ORF4和ORF5对WAP-8294A的生物合成都是必需的。为了进一步测试ORF4的功能,我们在体内构建了两个ORF4活性单点突变体,H104A或H298A,在这两个突变体中ORF4保持完整,只有一个氨基酸改变。HPLC和LC-MS分析结果表明,从单点突变株OH11WAP-ORF4-H104A或OH11WAP-ORF4-H298A代谢产物中均未检测到WAP-8294A化合物。ORF4预测为编码2-酮戊二酸依赖性双加氧酶,该酶催化WAP-8294A化合物中D-Asn残基的β羟基化或脂肪酸部分的β羟基化。为了进一步检测单点突变株的代谢物,我们进行了高分辨率LC-MS分析,并与野生型的代谢物进行了比较。结果表明ORF4单点突变体能够产生新的WAP-8294A类似物,其缺少母体WAP-8294A化合物的羟基。该结果为ORF4在WAP-8294A生物合成中D-Asn残基的β-羟基化中的功能提供了证据。随后我们对OH11野生型UHPLC-HR-MS/MS图谱中主峰附近的6个小峰进行了分析,鉴定结果表明它们均与骨架氨基酸略有不同(AZ1、AZ2、AZ3 和AZ6)或者D-3-OH的脂肪酸链略有差异(AZ4)的WAP-8294A家族新成员(图4)。相同UHPLC-HR-MS/MS数据的分子网络分析结果也鉴定到了这6个新化合物(图3),有趣的是,AZ5似乎与先前报道的微量化合物WAP-8294Ax8相同。
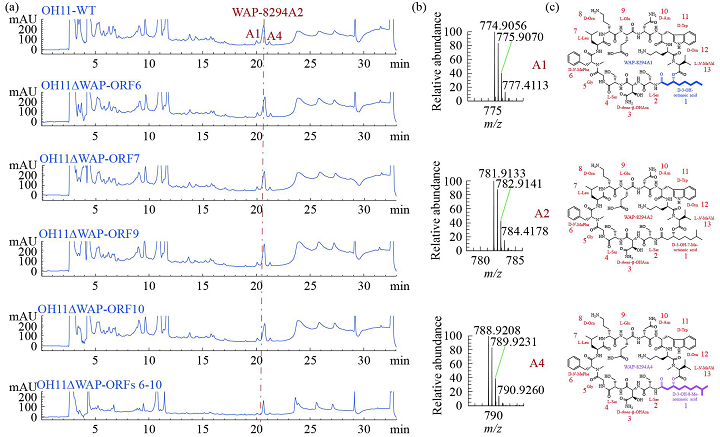
图2 UHPLC-HR-MS/MS分析产酶溶杆菌WAP基因簇中辅助基因同框缺失突变株代谢物
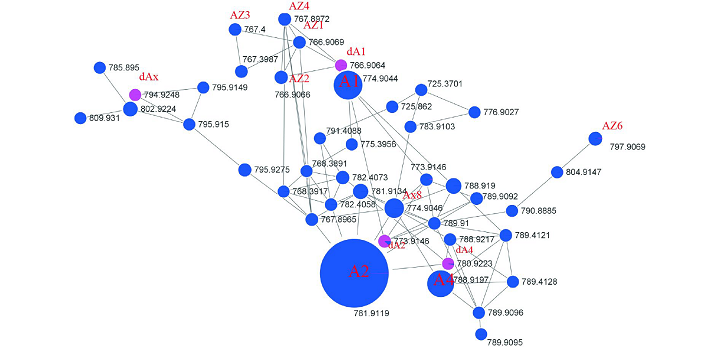
图3 突变株OH11ΔWAP-ORF4和野生型OH11中WAP-8294代谢物的分子网络分析
▎讨论
有研究报道了用化学法合成WAP-8294A2及其脱氧类似物脱氧WAP-8294A2,并证实了脱氧-WAP-8294A2也是有效的抗菌化合物。但是其合成过程需要27步高度复杂的化学反应,因此通过化学合成方法获得大量该化合物可行性不高,如果能将突变株产量提高,生物合成方法可能是一种可行的替代方法。
在本研究中,我们使用UHPLC-HR-MS/MS分析产酶溶杆菌OH11的代谢物,并鉴定了六种新化合物WAP-8294AZ1、AZ2、AZ3、AZ4、AZ6和AZ7,以及四种已知化合物A1、A2、A4和Ax8(AZ5)(图4)。新化合物包含氨基酸残基的变化(AZ1、AZ2、AZ3和AZ6)或D-3-OH脂肪酸的变化(AZ4)(图1)。最值得注意的是,AZ7仅含有A2的9个氨基酸残基(图4)。这9个氨基酸残基的组成和序列与WAP-8294A2的第4–12个残基相同。这表明AZ7可能和溶杆菌3655中lysocins的降解情况一样,是母体化合物的降解产物,或者AZ7是异常从4位起始12位释放并环化的反常规生物合成过程的产物,这种情况在溶杆菌的NRPS种也有过报道。
发现能生产具有新的化学结构和作用机理的天然产物的生防菌可以减少使用化学农药带来的环境的问题,为解决病原菌抗药性问题提供绿色高效的策略。本研究表明,可以在不引入外源DNA并对溶杆菌生物合成基因进行最低限度的遗传操作便可以产生新的抗菌WAP-8294A化合物。本工作为今后改造WAP生物合成途径增加新化合物产量,以及为将溶杆菌开发成管理作物病害的安全有效的工具奠定了基础。
· 文 章 链 接 ·
 返回列表
返回列表